This opinion paper is inspired
by scientific discussion of our presentation at the National Institutes of
Health Conference "The Dynamic and Energetic Bases of Health and
Aging"(NIH,
We are grateful to Drs. Bortz, Yates, Martin, Sing, and other participants of this
NIH conference for encouraging and useful discussion of ideas presented at this
Confetence.
March 9,
The Quest
for the General Theory
of Aging and Longevity
Leonid A. Gavrilov*,
Natalia S. Gavrilova
Center on Aging,
NORC/University of
* Corresponding author
E-mail address: gavrilov@aol.com (L.A.Gavrilov).
website: http://longevity-science.org
Address for correspondence:
Dr. Leonid
A. Gavrilov, Center on Aging
NORC/University of
Fax: (773) 256-6313; Phone:
(773) 256-6359
Extensive studies of
aging phenomenon have produced many important and diverse findings, which
require a general theoretical framework for them to be organized into a
comprehensive body of knowledge.
As demonstrated by the
success of evolutionary theories of aging, based on a general idea of the
declining force of natural selection with age, quite general theoretical
considerations can in fact be very useful and practical when applied to aging
research.
In this study, we
attempt to go one step further in the search for a general explanation of aging
by applying a general theory of systems failure known as reliability theory.
Considerations of this
theory lead to the following conclusions:
(1) Redundancy is
a key notion for understanding aging and the systemic nature of aging in
particular. Systems, which are redundant in numbers of irreplaceable elements,
do deteriorate (i.e., age) over time, even if they are built of non-aging
elements.
(2) An apparent aging
rate or expression of aging (measured as age differences in failure rates,
including death rates) is higher for systems with higher redundancy levels.
(3) Redundancy
exhaustion over the life course explains the observed ‘compensation law of
mortality’ (mortality convergence at later life) as well as the observed
late-life mortality deceleration, leveling-off, and mortality plateaus.
(4) Living organisms
seem to be formed with a high load of initial damage, and therefore
their lifespans and aging patterns may be sensitive
to early-life conditions that determine this initial damage load during
early development. The idea of early-life programming of aging and longevity
may have important practical implications for developing early-life
interventions promoting health and longevity.
The theory also suggests
that aging research should not be limited to the studies of qualitative changes
(like age changes in gene expression), because changes in quantity
(numbers of cells and other functional elements) could be an important driving
force of aging process. In other words, aging may be largely driven by a
process of redundancy loss.
Finally we suggest
adding the reliability theory to the arsenal of methodological approaches
applied in aging research.
1. Introduction
There is a growing
interest in scientific explanations of aging and in the search for a general
theory that can explain what aging is and why and how it happens.
There is also a need for
a general theoretical framework that would allow researchers to handle an
enormous amount of diverse observations related to aging phenomena. Empirical
observations on aging have become so abundant that a special 4-volume
encyclopedia, The Encyclopedia of Aging, is now required for even
partial coverage of the accumulated facts (Ekerdt,
2002). To transform these numerous and diverse observations into a
comprehensive body of knowledge, a general theory of species aging and
longevity is required.
The prevailing research
strategy now is to focus on the molecular level in the hope of understanding
the proverbial nuts and bolts of the aging process. In accordance with this
approach, many aging theories explain aging of organisms through aging of
organisms’ components. However, this circular reasoning of assuming aging in
order to "explain" aging leads to a logical contradiction, because
moving in succession from the aging of organisms to the aging of organs,
tissues, and cells, we eventually come to atoms, which are known not to age.
Thus we come to the
following basic question on the origin of aging: How can we explain the
aging of a system built of non-aging elements?
This question invites us
to start thinking about the possible systemic nature of aging and to wonder
whether aging may be a property of the system as a whole.
In other words, perhaps
we need to broaden our vision and be more concerned with the bigger picture of
the aging phenomenon rather than its tiny details.
To illustrate the need
for a broad vision, consider the following questions:
-- Would it be possible
to understand a newspaper article by looking at it through an electronic
microscope?
-- Would the perception
of a picture in an art gallery be deeper and more comprehensive at the shortest
possible distance from it?
Evolutionary perspective
on aging and longevity is one way to stay focused on the bigger picture (see
reviews in Le Bourg, 2001; Gavrilova and Gavrilov, 2002). Evolutionary explanations of aging
and limited longevity of biological species are based on two major evolutionary
theories: the mutation accumulation theory (Medawar,
1946) and the antagonistic pleiotropy theory
(Williams, 1957). These two theories can be summarized as follows:
(1) Mutation
accumulation theory: From the evolutionary perspective, aging is an inevitable
result of the declining force of natural selection with age. For example, a
mutant gene that kills young children will be strongly selected against (will
not be passed to the next generation) while a lethal mutation that affects only
people over the age of 80 will experience no selection because people with this
mutation will have already passed it to their offspring by that age. Over
successive generations, late-acting deleterious mutations will accumulate,
leading to an increase in mortality rates late in life.
(2) Antagonistic pleiotropy theory: Late-acting deleterious genes may even
be favored by selection and be actively accumulated in populations if they have
any beneficial effects early in life.
Note that these two
theories of aging are not mutually exclusive, and both evolutionary mechanisms
may operate at the same time. The main difference between the two theories is
that in the mutation accumulation theory, genes with negative effects at old
age accumulate passively from one generation to the next while in the
antagonistic pleiotropy theory,
these genes are actively kept in the gene pool by selection (Le Bourg, 2001).
The actual relative contribution of each evolutionary mechanism to species
aging has not yet been determined, and this scientific problem is now the main
focus of current research in evolutionary biology.
Evolutionary theories
demonstrate that taking a step back from too close consideration of the details
over "the nuts and bolts" of the aging process helps to gain a
broader vision of the aging problem.
The remaining question
is whether the evolutionary perspective represents the ultimate general
theoretical framework for explanations of aging. Or perhaps there may be even
more general theories of aging, one step further removed from the particular
details?
The main limitation of
evolutionary theories of aging is that they are most applicable to sexually
reproducing organisms, because these theories are based on the idea of natural
selection and on the declining force of natural selection with age.
However, aging is a very
general phenomenon -- it is also observed in technical devices (like cars),
which do not reproduce themselves in a sexual or any other way and which are,
therefore, not subject to evolution through natural selection.
Thus, there may exist a more general explanation of aging, beyond mutation
accumulation and antagonistic pleiotropy theories.
The quest for a general
explanation of aging (age-related increase in failure rates), applicable both
to technical devices and biological systems invites us to consider the general
theory of systems failure known as reliability theory (Gavrilov
and Gavrilova, 2001a).
2.
General Overview of Reliability Theory Approach
Reliability theory is a
body of ideas, mathematical models, and methods directed to predict, estimate,
understand, and optimize the lifespan distribution of systems and their
components (Barlow and Proschan, 1975; Gavrilov and Gavrilova,
2001a). The reliability of the system (or component) refers to its
ability to operate properly according to a specified standard (Crowder et al.,
1991). Reliability is described by the reliability function S(x), which
is the probability that a system (or component) will carry out its mission
through time x (Rigdon and Basu,
2000). The reliability function (also called the survival function)
evaluated at time x is simply the probability P, that the failure
time X, is beyond time x, designated as P(X > x).
Thus, the reliability function is represented in the following way:
![]()
where F(x) is a standard cumulative
distribution function from the probability theory (Feller, 1968). The best
illustration for the reliability function S(x) is a survival curve describing
the proportion of those still alive by time x (the lx
column in life tables). The failure rate l(x), also called the hazard rate h(x),
is defined as the relative rate for reliability function decline:
![]()
Failure rate is an
equivalent to mortality force, m(x), in demography and gerontology. When the
failure rate is constant (i.e., does not increase with age), we have a non-aging
system (component) that does not deteriorate (does not fail more often)
with age. The reliability function of non-aging systems (components) is
described by the exponential distribution:
![]()
This failure law
describes the 'lifespan' distribution of atoms of radioactive elements, and it
is also observed in many wild populations with high extrinsic mortality (Finch,
1990; Gavrilov and Gavrilova,
1991).
Non-aging behavior of a
system can be detected graphically, when the logarithm of the survival function
decreases with age in a linear fashion:
ln S(x) = ln S0
- λx (3c)
Interestingly, the
survival patterns of humans at extreme old ages (over 100 years old) are rather
close to this linear dependence, suggesting that death rates, although very
high, do not demonstrate further dramatic deterioration with age (Figure 1).
Figure
1 About Here
The same phenomenon
of 'almost non-aging' survival dynamics at extreme old ages is detected
in many other biological species including rodents (guinea pigs, rats, and mice)
and invertebrates (nematodes, shrimps, bdelloid
rotifers, Drosophila, Campanularia Flexuosa), – a phenomenon well known since the
1970s (Economos, 1979), but still presenting a
theoretical challenge to many gerontologists. One interesting corollary
from these intriguing observations is that there seems to be no fixed upper
limit for individual lifespan (Gavrilov, 1984; Gavrilov and Gavrilova, 1991).
Interestingly, the
failure kinetics of manufactured products (steel samples, industrial relays,
and motor heat insulators) also demonstrates the same 'non-aging' pattern at
the end of their 'lifespan' (Economos, 1979).
This observation calls for a very general explanation of this apparently
paradoxical 'no aging at extreme ages'
phenomenon, which will be suggested in this article.
If failure rate
increases with age, we have an aging system (component) that
deteriorates (fails more often) with age. There are many failure laws for aging
systems and the Gompertz law with
exponential increase of the failure rates with age (Gompertz,
1825) is just one of them (see Gavrilov and Gavrilova, 1991). In reality, system failure rates may
contain both non-aging and aging terms as, for example, in the case of the Gompertz-Makeham law of mortality (Makeham, 1860; Strehler, 1978; Gavrilov and Gavrilova, 1991):
![]()
In this formula the first,
age-independent term (Makeham parameter, A)
designates the constant, 'non-aging' component of the failure rate (presumably
due to extrinsic causes of death, such as accidents and acute infections),
while the second, age-dependent term (the Gompertz
function, R× eax) designates the 'aging' component, presumably
due to deaths from age-related degenerative diseases like cancer and heart
disease.
The validity of the Gompertz-Makeham law of mortality (4) can be illustrated
graphically, when the logarithms of death rates without the Makeham
parameter (μx - A) are
increasing with age in a linear fashion:
log
(μx - A) = log(R) +
α x
(5)
where log(R) is an intercept coefficient and α is a slope coefficient in this
linear relationship.
The log-linear increase
in death rates (adjusted for the Makeham term) with
age is indeed a very common phenomenon for many human populations at ages 35-70
years (see Figure 2).
Figure 2 About Here
Note that the slope
coefficient α characterizes an "apparent aging
rate" (how rapid is the age-deterioration in mortality) -- if α is equal to zero, there is no
apparent aging (death rates do not increase with age).
At advanced ages (after
age 70), the 'old-age mortality deceleration' takes place -- death rates are
increasing with age at a slower pace then expected from the Gompertz-Makeham
law. This mortality deceleration eventually produces "late-life mortality
leveling-off" and "late-life mortality plateaus" at extreme old
ages (Gavrilov and Gavrilova,
1991; 2001a).
The compensation law
of mortality in its strong form refers to mortality convergence,
when higher values for the parameter a (in the Gompertz function) are compensated for by lower values of
the parameter R in different populations of a given species:
![]()
where B and M are universal
species-specific invariants. Sometimes this relationship is also called the Strehler-Mildvan correlation (Strehler
and Mildvan, 1960; Strehler,
1978), although that particular correlation was largely an artifact of the
opposite biases in parameters’ estimation caused by not taking into account the
age-independent mortality component, the Makeham term
A (see Gavrilov and Gavrilova,
1991). Parameter B is called the species-specific lifespan (95 years for
humans), and parameter M is called the species-specific mortality rate (0.5
year-1 for humans). These parameters are the coordinates for
convergence of all the mortality trajectories into one single point (within a
given biological species), when extrapolated by the Gompertz
function (Figure 2).
In those cases when the
compensation law of mortality is not observed in its strong form, it may still
be valid in its weak form – i.e., the relative differences in mortality rates
of compared populations tend to decrease with age in many species. Explanation
of the compensation law of mortality is a great challenge for many theories of
aging and longevity (Strehler, 1978; Gavrilov and Gavrilova, 1991;
2001a).
There are some
exceptions both from the Gompertz law of mortality
and the compensation law of mortality that have to be understood and explained.
In some cases the organisms die according to the Weibull
(power) law (Hirsch et al., 1994; Eakin et al.,
1995; Vanfleteren et al., 1998):
m(x) =lxa for x ³ 0, where l, a > 0 (7)
The validity of the Weibull law can be illustrated graphically, when the
logarithm of the failure rate increases in a linear fashion as a function of
the logarithm of age:
log
[m(x)] =
log(l) + a Z, where Z = log x (8)
Here log(l) is an intercept coefficient and a a slope coefficient in this linear
relationship. Some examples of such linear dependence will be provided
later (Figure 3).
Note that the slope
coefficient a in the Weibull law characterizes an 'apparent aging rate' (the rapidity
of the age-deterioration in mortality), – if a is equal to zero, there is no
apparent aging (death rates do not increase with age).
The Weibull
law is more commonly applicable to technical devices (Weibull,
1951; Barlow and Proschan, 1975; Rigdon
and Basu, 2000), while the Gompertz
law is more common to biological systems (Strehler,
1978; Finch, 1990; Gavrilov and Gavrilova,
1991). Possible explanations why organisms prefer to die according to the
Gompertz law, while technical devices typically fail
according to the Weibull law are provided elsewhere (Gavrilov and Gavrilova, 1991;
2001a) and will be briefly discussed later.
The phenomena of
mortality increase with age and the subsequent mortality leveling-off are
theoretically predicted to be an inevitable feature of all reliability models
that consider aging as a progressive accumulation of random damage (Gavrilov and Gavrilova, 1991;
2001). Mathematical illustration for this statement is provided in the next
section of this paper. In simple words, if the destruction of an organism
occurs not in one but in two or more sequential random stages, this is
sufficient for the phenomenon of aging (mortality increase) to appear and then
to vanish at older ages. Each stage of destruction corresponds to one of the
organism's vitally important structures being damaged. In the simplest
organisms with unique, critical structures, this damage usually leads to
death. Therefore defects in such organisms do not accumulate, and the
organisms themselves do not age – they just die when damaged. In more complex
organisms with many vital structures and significant redundancy, every
occurrence of damage does not lead to death because of this redundancy. Defects
do accumulate, therefore, giving rise to the phenomenon of aging (mortality
increase). Thus, aging is a direct consequence (trade-off) of systems’
redundancies that ensure increased reliability and increased lifespan of
organisms. As defects accumulate, the redundancy in the number of elements
finally disappears. As a result of this redundancy exhaustion, the organism
degenerates into a system with no redundancy, that is, a system with elements
connected in series, with the result being that any new defect leads to death.
In such a state, no further accumulation of damage can be achieved, and the
mortality rate levels off. The next section provides mathematical illustration
for these ideas.
3. Explanations of Aging Phenomena Using Reliability Theory:
An Illustrative Example
Consider a system built of non-aging
elements with a constant failure rate k. If these n elements are
mutually substitutable, so that the failure of a system occurs only when all
the elements fail (parallel construction in the reliability theory context),
the cumulative distribution function for system failure, F(n,k,x), depends on age x in the
following way:
![]()
Therefore,
the reliability function of a block, Sb(n,k,x) can be represented as:
![]()
Consequently,
the failure rate of a block mb(n,k,x) can be written as follows:
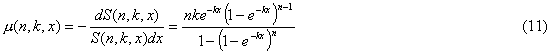
» nknxn-1 (11a)
when x << 1/k (early-life period approximation, when 1-e-kx
» kx);
» k
(11b)
when x >> 1/k (late-life period approximation, when 1-e-kx
» 1)
Thus, the failure rate
of a system initially grows as a power function of age (the Weibull
law). Then the tempo at which the failure rate grows declines,
and the failure rate approaches asymptotically an upper limit equal to k.
Here we should pay attention to three significant points. First, a system
constructed of non-aging elements is now behaving like an aging object: i.e.,
aging is a direct consequence of the redundancy of the system (redundancy in
the number of elements). Second, at very high ages the phenomenon of aging
apparently disappears (failure rate levels-off), as redundancy in the number of
elements vanishes. The failure rate approaches an upper limit, which is totally
independent of the initial number of elements, but coincides with the rate of
their loss (parameter k). Third, the systems with different initial
levels of redundancy (parameter n) will have very different failure
rates in early life, but these differences will eventually vanish as failure
rates approach the upper limit determined by the rate of elements' loss
(parameter k). Thus, the compensation law of mortality (in its weak
form) is an expected outcome of this illustrative model. These
theoretical predictions are supported by experimental studies on Drosophila melanogaster, which found no differences in late-life
mortality between cohorts of flies having markedly different levels of early
robustness (Drapeau et al., 2000).
These theoretical
statements, based on general analytical considerations, are also illustrated
here with the following particular numerical example. Fig. 3 presents the
results of computer simulation of mortality kinetics in systems with different
levels of redundancy. Specifically, calculations of failure rates are
performed according to formula (11) described earlier, for the numbers of
elements, n = 1, 2, 3, 4, and 5. The scales for mortality rates (vertical
axis), and for age (horizontal axis) are presented in dimensionless units (μ/k
for mortality rates, and kx for age), to
ensure the generalizability of the results
(invariance of graphs on failure rate of the elements in the system, parameter k).
Also, the log scale is used to explore the system behavior in a wide range of
ages (0.01 - 10 units), and failure rates ( 0.00000001
- 1.0 units).
Figure 3 About Here
This graph depicts mortality trajectories for five
systems with different degrees of redundancy:
(1) System # 1 has only one unique element (no
redundancy), and it has the highest failure rate, which does not depend on age
(no aging).
(2) System # 2 has two elements connected in parallel
(one extra element is redundant), and the failure rate is initially increasing
with age (aging appears). The apparent rate of aging can be characterized by a
slope coefficient, which is equal to one. Finally the failure rate levels-off
at advanced ages.
(3) System # 3 has three elements connected in parallel
(two extra elements are redundant), and the failure rate is initially
increasing with age (an apparent aging rate, a slope coefficient is equal to
two). Then the failure rate levels-off at advanced ages.
(4) System # 4 has four elements connected in parallel
(three extra elements are redundant, degree of redundancy = 3), and the failure
rate is initially increasing with age with slope coefficient = 3 (apparent
relative aging rate). Then again the failure rate levels-off at advanced
ages.
(5) System # 5 has five elements connected in parallel
(four extra elements are redundant, degree of redundancy = 4), and the failure
rate is initially increasing with age with the steepest slope coefficient = 4
(apparent relative aging rate). Finally, the mortality trajectory
levels-off at advanced ages.
This computational example illustrates the following
statements:
(1) Aging is a direct consequence of a system's
redundancy, and the expression of aging is directly related to the degree of a
system's redundancy. Specifically, an apparent relative aging rate is just
equal to a degree of redundancy in parallel systems.
(2) All mortality trajectories tend to converge with
age, so that the compensation law of mortality is observed.
(3) All mortality trajectories level-off at advanced
ages, and a mortality plateau is observed.
Thus, the major aging phenomena (aging itself, the
compensation law of mortality, late-life mortality deceleration, and late-life
mortality plateaus) are already observed in the simplest redundant systems.
However, to explain the Gompertz law of mortality, an
additional idea should be taken into account (see later).
4. The Idea of High Initial Damage Load
Reliability theory
predicts that a failure rate of simple redundant systems increases with age
according to the Weibull (power) law (see formula
11a, and Figure 3). This theoretical prediction is consistent with empirical
observations that failure kinetics of technical devices follows the Weibull law (Weibull, 1951; Gavrilov and Gavrilova, 1991,
2001a). However, biological systems 'prefer' to fail according to the Gompertz (exponential) law (Gompertz,
1825; Finch, 1990; Gavrilov and Gavrilova,
1991, 2001a), which calls for explanations.
An attempt to explain
exponential deterioration of biosystems in terms of
the reliability theory had led to a paradoxical conjecture that biological
systems start their adult life with high load of initial damage (Gavrilov and Gavrilova, 1991,
2001a).
Although this idea may
look like a counter-intuitive assumption, it fits well with many empirical
observations on massive cell losses in early development. For example,
the female human fetus at age 4-5 months possesses 6-7 million eggs (oocytes). By birth, this number drops to 1-2 million and
declines even further. At the start of puberty in normal girls, there are
only 0.3-0.5 million eggs – just only 4-8% of initial numbers (Finch and
Massive cell losses in
early development are creating conditions for Poisson distribution of organisms
according to the numbers of remaining cells, which in turn produce the
exponential (Gompertzian) law of mortality increase (Gavrilov and Gavrilova, 1991).
Because the mathematical proof for this statement is already published
elsewhere (Gavrilov and Gavrilova,
1991), we can concentrate here on substantive discussion of the idea of high
initial damage load in biological systems.
Biological systems are different
from technical devices in two aspects. The first fundamental feature of biosystems is that, in contrast to technical (artificial)
devices which are constructed out of previously manufactured and tested
components, organisms form themselves in ontogenesis through a process of
self-assembly out of de novo forming and externally untested elements (cells).
The second property of organisms is the extraordinary degree of miniaturization
of their components (the microscopic dimensions of cells, as well as the
molecular dimensions of information carriers like DNA and RNA), permitting the
creation of a huge redundancy in the number of elements. Thus, we can expect
that for living organisms, in distinction to many technical (manufactured)
devices, the reliability of the system is achieved not by the high initial
quality of all the elements but by their huge numbers (redundancy).
The fundamental difference in the
manner in which the system is formed (external assembly in the case of
technical devices and self-assembly in the case of biosystems)
has two important consequences. First, it leads to the macroscopicity
of technical devices in comparison with biosystems,
since technical devices are assembled 'top-down' with the participation of a
macroscopic system (man) and must be suitable for this macroscopic system to
use (i.e., commensurate with man). Organisms, on the other hand, are assembled
'bottom-up' from molecules and cells, resulting in an exceptionally high degree
of miniaturization of the component parts. Second, since technical devices are
assembled under the control of man, the opportunities to pretest components (external
quality control) are incomparably greater than in the self-assembly of biosystems. The latter inevitably leads to organisms being
'littered' with a great number of defective elements. As a result, the
reliability of technical devices is assured by the high quality of elements,
with a strict limit on their numbers because of size and cost limitations,
while the reliability of biosystems is assured by an
exceptionally high degree of redundancy to overcome the poor quality of some
elements.
It is interesting to note that the
uniqueness of individuals, which delights biologists so much, may be caused by
'littering' the organisms with defects and thus forming a unique pattern of
individual damage. Our early experience working with dilapidated computer equipment
in
The idea that living
organisms are starting their lives with a large number of defects has deep
historical roots. Biological justification for this idea was discussed by Dobzhansky (1962). He noted that, from the biological
perspective, Hamlet’s “thousand natural shocks that flesh is heir to”
was an underestimate and that in reality “the shocks are innumerable” (Dobzhansky, 1962, p.126). Recent studies found that the
troubles in human life start from the very beginning: the cell-cycle
checkpoints (which ensure that cell will not divide until DNA damage is
repaired and chromosomal segregation is complete) do not operate properly at
early, cleavage stage of human embryo (Handyside and Delhanty, 1997). This produces mosaicism
of the preimplantation embryo, where some embryonic
cells are genetically abnormal (McLaren, 1998) with
potentially devastating consequences in later life. Most of the DNA
damage caused by copy-errors during DNA replication also occurs in early life,
because most cell divisions happen in early development. As a result of extensive DNA damage in early development, many
apparently normal tissues of young organisms have an unbelievably high load of
mutations, including an amazing amount of oncogenic
mutations and frequent clones of mutated somatic cells (Cha et. al., 1994; Deng
et al., 1996; Johason et al., 1996).
Another potential source
of extensive initial damage is the birth process itself. During birth, the
future child is first deprived of oxygen by compression of the umbilical cord
(Moffett et al., 1993), and suffers severe hypoxia (often with ischemia
and asphyxia). Then, just after birth, a newborn child is exposed to oxidative
stress because of acute reoxygenation while starting
breathing. It is known that acute reoxygenation after
hypoxia may produce an extensive oxidative damage through the same mechanisms
that also produce ishemia-reperfusion injury (IRI)
and asphyxia-reventilation injury (Martin et al.,
2000). Thus, using Hamlet's metaphor, we may conclude that humans "suffer
the slings and arrows of outrageous fortune" and have "a sea
of troubles" from the very beginning of their lives.
It follows from this concept of high
initial damage load that even small progress in optimizing the processes of
ontogenesis and increasing the numbers of initially functional elements can
potentially result in a remarkable fall in mortality and a significant
improvement in lifespan. This optimistic prediction is supported by
experimental evidence of increased offspring lifespan in response to protection
of parental germ cells against oxidative damage just by feeding the future
parents with antioxidants (Harman and Eddy, 1979). Increased lifespan is also
observed among the progeny of parents with a low respiration rate (proxy for
the rate of oxidative damage to DNA of germ cells, see Gavrilov
and Gavrilova, 1991). The concept of high
initial damage load also predicts that early life events may affect survival in
later adult life through the level of initial damage. This prediction proved to
be correct for such early life indicators as parental age at a person's
conception (Gavrilov and Gavrilova,
1997a; 1997b; 2000; 2001b; 2003) and the month of person's birth (Gavrilov and Gavrilova, 1999;
2003; Doblhammer and Vaupel,
2001). There is mounting evidence now in support of the idea of fetal origins
of adult degenerative diseases (Barker, 1998; Kuh and
Ben-Shlomo, 1997; Leon et al., 1998; Lucas et al.,
1999), and early-life programming of aging and longevity (Gavrilov
and Gavrilova, 1991; 2001b; 2003).
5.
Concluding Remarks
Extensive studies in
experimental gerontology have produced many important and diverse findings,
which require a general theoretical framework for them to be organized into a
comprehensive body of knowledge.
As demonstrated by the
success of evolutionary theories of aging, based on a general idea of the
declining force of natural selection with age, quite general theoretical
considerations can in fact be very useful and practical when applied to aging
research (Le Bourg, 2001; Gavrilov and Gavrilova, 2002).
In this study, we attempted
to go one step further in the search for a general explanation of aging by
applying a general theory of systems failure known as reliability theory.
Considerations of this
theory lead to the following conclusions:
(1) Redundancy is
a key notion for understanding aging and the systemic nature of aging in
particular. Systems, which are redundant in numbers of irreplaceable elements,
do deteriorate (i.e., age) over time, even if they are built of non-aging
elements. The positive effect of systems’ redundancy is damage
tolerance, which decreases mortality and increases lifespan. However
damage tolerance makes it possible for damage to be tolerated and accumulated
over time, thus producing aging phenomenon.
(2) An apparent aging
rate or expression of aging (measured as age differences in failure rates,
including death rates) is higher for systems with higher redundancy levels (all
other things being equal). This is an important issue, because it helps
to put a correct perspective over fascinating observations of negligible
senescence (no apparent aging) observed in the wild and at extreme old ages.
Reliability theory explains that some cases of negligible senescence may have a
trivial mechanism (lack of redundancies in the system being exposed to challenging
environment) and, therefore, will not help to uncover "the secrets of
negligible senescence". The studies of negligible senescence make
sense however when the total mortality rates are also demonstrated to be
negligible.
(3) During the life
course the organisms are running out of their cells (Finch and
(4) Living organisms
seem to be formed with a high load of initial damage, and therefore
their lifespans and aging patterns may be sensitive
to early-life conditions that determine this initial damage load during
early development. The idea of early-life programming of aging and longevity
may have important practical implications for developing early-life
interventions promoting health and longevity.
The theory also suggests
that aging research should not be limited to the studies of qualitative changes
(like age changes in gene expression), because changes in quantity
(numbers of cells and other functional elements) could be an important driving
force of aging process. In other words, aging may be largely driven by a process
of redundancy loss.
The reliability theory
predicts that a system may deteriorate with age even if it is built from
non-aging elements with constant failure rate. The key issue here is the
system's redundancy for irreplaceable elements, which is responsible for the
aging phenomenon. In other words, each particular step of system
destruction/deterioration may seem to be apparently random (no aging, just
occasional failure by chance), but if a system failure requires a sequence of
several such steps (not just a single step of destruction),
then system as a whole may have an aging behavior.
Why is this important?
Because the significance of beneficial health-promoting interventions is often
undermined by claims that these interventions are not proven to delay the
process of aging itself, but instead that they simply delay or
"cover-up" some particular manifestations of aging.
In contrast to these
pessimistic views, the reliability theory says that there may be no specific
underlying elementary "aging process itself" – instead aging may be
largely a property of redundant system as a whole, because it has a network of
destruction pathways each being associated with particular manifestations of
aging (types of failure). Therefore, we should not be discouraged by only
partial success of each particular intervention, but instead we can appreciate
an idea that we do have so many opportunities to oppose aging in numerous
different ways.
Thus, the efforts to
understand the routes and the early stages of age-related degenerative diseases
should not be discarded as irrelevant to understanding of the "true
biological aging". On the contrary, the attempts to build an intellectual
firewall between experimental gerontology and clinical medicine are
counterproductive. After all, the main reason why people are really concerned
about aging is because it is related to health deterioration and increased
morbidity. The most important pathways of age changes are those that make older
people sick.
Ageing is a complex phenomenon and a holistic
approach using reliability theory may help to analyze, understand and perhaps
to control it.
Finally we suggest
adding the reliability theory to the arsenal of methodological approaches
applied in aging research.
Acknowledgments
We would like to thank participants
of three scientific meetings where the ideas and findings of this study were
presented and discussed:
(1) Annual meeting of
the Gerontological Society of America (
(2) National Institutes
of Health Conference "The Dynamic and Energetic Bases of Health and
Aging"(NIH,
(3) Annual Meeting of
the Population Association of
We are most grateful to
Mr. Brian Whiteley for useful editorial
suggestions. We would also like to acknowledge partial support from the
National Institute on Aging grants.
References
Barker, D.J.P. 1998. Mothers, Babies, and Disease in Later Life. 2nd edition. Churchill Livingstone,
Barlow, R.E.
and Proschan, F. 1975. Statistical
Theory of Reliability and Life Testing. Probability
Models. Holt, Rinehart and
Cha, R.S., Thilly,
W.G., Zarbl, H., 1994. N-nitroso-N-methylurea-induced
rat mammary tumors arise from cells with preexisting oncogenic
Hras1 gene mutations. Proc. Natl. Acad. Sci.
USA 91, 3749-3753.
Crowder, M.J., Kimber, A.C., Smith, R.L., Sweeting, T.J., 1991. Statistical Analysis of
Reliability Data. Chapman & Hall,
Deng, G., Lu, Y., Zlotnikov, G., Thor, A. D., Smith, H. S., 1996. Loss of heterozygosity in normal tissue
adjacent to breast carcinomas. Science 274, 2057-2059
Doblhammer, G., Vaupel, J.W.,
2001. Lifespan depends on month of birth. Proc. Natl. Acad.
USA 98, 2934-2939.
Dobzhansky, T., 1962. Mankind Evolving.
The Evolution of Human Species.
Drapeau,
M.D., Gass, E.K., Simison,
M.D., Mueller, L.D., Rose, M.R., 2000. Testing the heterogeneity theory of
late-life mortality plateaus by using cohorts of Drosophila melanogaster. Exp. Gerontol.
35, 71-84.
Eakin, T., Shouman,
R., Qi, Y.L., Liu, G.X.,
Economos,
A.C., 1979. A non-gompertzian paradigm for mortality
kinetics of metazoan animals and failure kinetics of manufactured products. AGE 2, 74-76.
Ekerdt,
D.J. (Ed.), 2002. The Encyclopedia of Aging. Macmillan
Reference
Feller, W., 1968. An Introduction to Probability Theory and its
Applications. Vol.1, Wiley and Sons,
Finch, C.E., 1990. Longevity, Senescence and the Genome.
Finch, C.E.,
Gavrilov L.A., 1984. Does a limit of the
life span really exist? Biophysics [Biofizika] 29,
908-911.
Gavrilov,
Gavrilov,
Gavrilov,
Gavrilov,
Gavrilov,
Gavrilov,
Gavrilov,
Gavrilov,
Gavrilov,
Gompertz,
B., 1825. On the nature of the function expressive of
the law of human mortality and on a new mode of determining life contingencies.
Philos.Trans.Roy.Soc.London A, 115, 513-585.
Handyside,
A.H., Delhanty, J.D.A., 1997. Preimplantation
genetic diagnosis: strategies and surprises. Trends in Genetics 13, 270-275.
Harman, D.,
Eddy, D.E., 1979. Free radical theory of aging: beneficial effects of adding
antioxidants to the maternal mouse diet on life span of offspring: possible
explanation of the sex difference in longevity. AGE 2,
109-122.
Hirsch,
A.G., Williams, R.J., Mehl, P., 1994. Kinetics of medfly mortality. Exp.
Gerontol. 29, 197-204.
Jonason, A. S., Kunala,
S., Price, G. T., Restifo, R. J., Spinelli,
H. M., Persing, J. A., Leffell,
D. J., Tarone, R. E., Brash, D. E., 1996. Frequent clones of p53-mutated keratinocytes
in normal human skin. Proc. Natl. Acad. Sci.
USA 93, 14025-14029.
Kuh, D., Ben-Shlomo,
B., 1997. A Life Course Approach to Chronic Disease Epidemiology.
Le Bourg, É., 2001. A mini-review of the
evolutionary theories of aging. Is it the time to accept them?
Demographic Research (online), 4(1), 1-28. Available from URL:
http://www.demographic-research.org/volumes/vol4/1/4-1.pdf
Lucas, A., Fewtrell, M. S., Cole, T. J., 1999. Fetal origins of adult
disease - the hypothesis revisited. Br. Med. J. 319, 245-249.
Makeham,
W.M., 1860. On the law of mortality and the construction of
annuity tables. J. Inst. Actuaries 8, 301-310.
Martin, L.J., Brambrink, A.M., Price, A.C.,
Kaiser, A., Agnew, D.M., Ichord, R.N., Traystman, R.J., 2000. Neuronal death in newborn striatum after
hypoxia-ischemia is necrosis and evolves with oxidative stress. Neurobiology of
Disease 7, 169-191.
McLaren,
A., 1998. Genetics and human reproduction. Trends in
Genetics 14, 427-431.
Medawar,
P.B., 1946. Old age and natural death. Mod.Q. 2, 30-49. [Reprinted in The
Uniqueness of the Individual, by P. B. Medawar, Basic
Books,
Moffett,
D.F., Moffett, S.B., Schauf, C.L., 1993. Human
Physiology: Foundations & Frontiers. 2nd ed., Wm. C.
Brown Publishers, Dubuque etc.
Rigdon,
S.E., Basu, A.P., 2000. Statistical
Methods for the Reliability of Repairable Systems. John Wiley &
Sons, Inc.,
Strehler,
B.L., 1978. Time, Cells, and Aging. 2nd
edition, Academic Press,
Strehler,
B.L., Mildvan, A.S., 1960. General
theory of mortality and aging. Science 132, 14-21.
Vanfleteren,
J.R., De Vreese, A., Braeckman,
B.P., 1998. Two-parameter logistic and Weibull
equations provide better fits to survival data from isogenic
populations of Caenorhabditis elegans
in axenic culture than does the Gompertz
model. J. Gerontol. Ser.
A 53, B393-403.
Weibull, W.A., 1951. A
statistical distribution function of wide applicability. J. Appl. Mech. 18, 293-297.
Williams,
G.C., 1957. Pleiotropy, natural selection and the evolution of senescence.
Evolution 11, 398-411.
Figure Captions
Figure 1.
Survival Patterns After
Age 90.
Percent surviving (in log scale) is plotted
as a function of age of Swedish women for calendar years 1900, 1980, and 1999
(cross-sectional data). Note that after age 100, the logarithm of
survival fraction is decreasing without much further acceleration (aging) in
almost a linear fashion. Also note an increasing pace of survival
improvement in history: it took less than 20 years (from year 1980 to year
1999) to repeat essentially the same survival improvement that initially took
80 years (from year 1900 to year 1980).
Source: cross-sectional (period)
life tables at the Berkeley Mortality Database (BMD):
http://www.demog.berkeley.edu/~bmd/
Figure 2.
Compensation Law of Mortality.
Convergence of
Mortality Rates (in different populations) at Advanced Ages.
Death rates (with removed Makeham parameter A,
corresponding to age-independent mortality component) are plotted in a log
scale as a function of age in the following countries:
1 –
2 –
3 –
4 - Northern Ireland, 1950-1952, males
5 -
6 -
7 -
Adapted from (Gavrilov et al., 1978; Gavrilov & Gavrilova, 1991)
Figure 3.
Failure kinetics of systems
with different levels of redundancy.
The dependence of the
logarithm of mortality force (failure rate) on logarithm of age in five systems
with different levels of redundancy (computer simulation experiment). Both the failure rate and the age are presented in
dimensionless units as explained in the text.
The dependence 1 is for the system containing only one
unique element (no redundancy). The dependence 2 is for the system containing
two elements connected in parallel (degree of redundancy = 1). The dependencies
3, 4 and 5 are for systems containing respectively 3, 4 and 5 elements
connected in parallel (with increasing levels of redundancy). Note that even in
this most simple case the following aging-related phenomena are observed: (1)
the emergence of aging as the system becomes redundant; (2) the increase in
apparent aging rate with increasing levels of system redundancy; (3) the
compensation law of mortality (mortality convergence), and (4) late-life
mortality deceleration and levelling-off to mortality
plateau. Additional explanations and comments are provided in the text of the
paper.
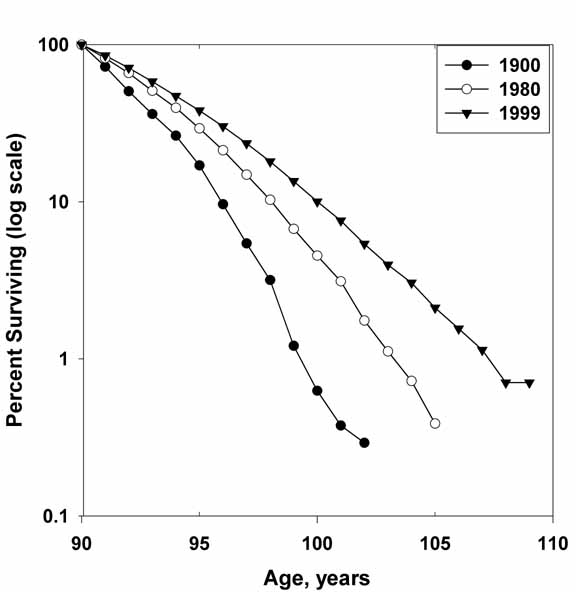
Figure 1
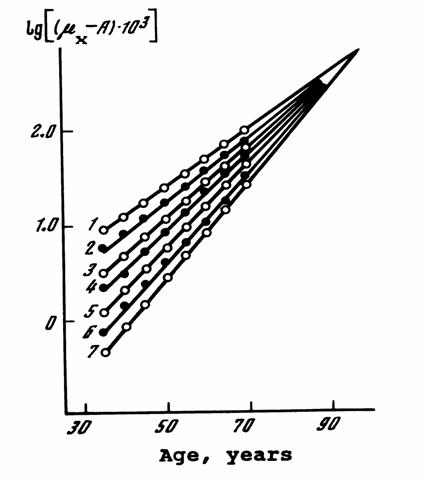
|
|
|
|
|
|
Figure 2
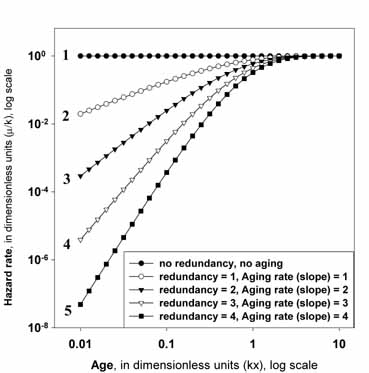
Figure 3